Volume 12 Issue 2 pp. 359-374 • doi: 10.15627/jd.2025.23
EEG-Based Neurophysiological Responses to Classroom Window Views in Green Campus Settings
Floriberta Binarti,* Nimas Sekarlangi, Meita Kasianus Virgin Brilianto, Bhanu Rizfa Hakim
Author affiliations
Department of Architecture, Universitas Atma Jaya Yogyakarta, Jl. Babarsari 44 Sleman 55281, Indonesia
*Corresponding author.
floriberta.binarti@uajy.ac.id (F. Binarti)
nimas.sekarlangit@uajy.ac.id (N. Sekarlangi)
200118133@students.uajy.ac.id (M. K. V. Brilianto)
239019746@students.uajy.ac.id (B. R. Hakim)
History: Received 24 April 2025 | Revised 16 June 2025 | Accepted 3 July 2025 | Published online 13 September 2025
2383-8701/© 2025 The Author(s). Published by solarlits.com. This is an open access article distributed under the terms and conditions of the Creative Commons Attribution 4.0 License.
Citation: Floriberta Binarti, Nimas Sekarlangi, Meita Kasianus Virgin Brilianto, Bhanu Rizfa Hakim, EEG-Based Neurophysiological Responses to Classroom Window Views in Green Campus Settings, Journal of Daylighting, 12:2 (2025) 359-374. doi: 10.15627/jd.2025.23
Figures and tables
Abstract
This study examines the neurophysiological responses of students to different classroom window views - forest, park, and city - within energy-efficient, green campus environments. Ten architecture students participated in EEG recordings while experiencing six virtual reality-simulated classroom conditions featuring varied window orientations and external views. The classroom models were designed to meet green building standards, with energy and daylighting performance validated using OTTV calculations and simulations in EnergyPlus and Radiance via DesignBuilder. EEG data were recorded using a 14-channel Emotiv EPOC X headset and analysed across delta to gamma frequency bands through power spectral and topographic mapping. Results revealed that forest views consistently evoked the most restorative neural responses, indicated by increased alpha and theta activity and decreased beta and gamma power. City views triggered neural patterns associated with cognitive load and overstimulation, while park views supported balanced attentional engagement. These findings highlight the importance of integrating biophilic elements - particularly dense greenery - into classroom design to enhance students’ cognitive performance and psychological well-being in sustainable educational settings.
Keywords
classroom window views, EEG brain activity, green campus environments, student well-being
Nomenclature
| Α | Solar absorptance |
| ΔT | Design temperature difference between outdoor and indoor (K) |
| OTTV | Overall Thermal Transfer Value (W/m²) |
| SC | Shading coefficient |
| SF | Solar factor (W/m²) |
| TDEk | Equivalent temperature difference (K) |
| Uf | Thermal transmittance of fenestration (W/m²·K) |
| Uw | Thermal transmittance of opaque wall (W/m²·K) |
Abbreviation
| AR | Augmented Reality |
| ART | Attention Restoration Theory |
| DA | Daylight Autonomy |
| ΔT | Temperature Difference |
| ECG | Electrocardiography |
| EEG | Electroencephalography |
| EMG | Electromyography |
| EN | European Norm (as in EN17037) |
| EOG | Electrooculography |
| ERP | Event-Related Potential |
| fMRI | Functional Magnetic Resonance Imaging |
| GBCI | Green Building Council Indonesia |
| HRV | Heart Rate Variability |
| ICA | Independent Component Analysis |
| LEED | Leadership in Energy and Environmental Design |
| LPP | Late Positive Potential |
| MR | Mixed Reality |
| OTTV | Overall Thermal Transfer Value |
| PSD | Power Spectral Density |
| SC | Shading Coefficient |
| SF | Solar Factor |
| SHGC | Solar Heat Gain Coefficient |
| TBR | Theta/Beta Ratio |
| TDEk | Equivalent Temperature Difference |
| Uf | Fenestration Thermal Transmittance |
| Uw | Wall Thermal Transmittance |
| VR | Virtual Reality |
| WWR | Window-to-Wall Ratio |
1. Introduction
Classroom environments play a critical role in supporting both instructional activities and students’ cognitive engagement. Research has consistently shown that the physical characteristics of classrooms significantly affect student satisfaction, attentional focus, and academic performance [1-5]. Within the paradigm of sustainable architecture, classroom windows are not only essential for optimizing energy efficiency but also function as key elements in cultivating health-promoting and conducive learning environments.
Properly designed windows contribute to improved indoor air quality, reduced energy usage, and enhanced visual and thermal comfort - all of which are closely linked to better student outcomes [6,7]. The visual connection to outdoor environments, especially natural scenes, has increasingly been recognized as an important factor in promoting student well-being. Exposure to natural landscapes - particularly greenery - has been associated with reduced stress, attentional recovery, better behavior, and enhanced academic performance [3,8-11]. Additionally, high-quality window views have been shown to elevate user satisfaction, mitigate visual discomfort, and increase tolerance to glare in both learning and working settings [12].
While much of the literature on window views has centered on office contexts, their significance in educational environments, particularly within green building frameworks, is gaining momentum. For example, [8] developed a computational tool using Seemo-survey data to assess view-related attributes, and [10] proposed a new method to quantify visual access to outdoor greenery. A survey-based study by [9] examined the relationship between the perceived naturalness of classroom window views and students’ well-being and academic outcomes, while [11] evaluated how nature visibility at school settings influenced behavioral issues. Similarly, Jiang, Li, Yongga, and Yan explored university students’ perceptions of classroom views, although their findings were limited due to constraints in window design and dimensions [13].
Studies investigating the influence of spatial attributes on human performance, behavior, and well-being fall under the emerging field of neuroarchitecture, which integrates four interconnected domains: neuroscience, architecture, physiology, and psychology (emotions and behavior) [14]. As noted by [14]. one of the goals of neuroarchitecture is to design environments that positively stimulate mental activity and influence psychological and physiological responses while minimizing stress. Attention Restoration Theory (ART) further supports this notion by suggesting that natural environments - key features in biophilic design - offer psychological refuge, promoting feelings of calm, safety, and well-being [15]. ART identifies core restorative qualities such as fascination, a sense of escape (being away), coherence or legibility of the environment (extent), and alignment between individual needs and environmental offerings (compatibility) [16].
A review by [12] on visual perception emphasized the importance of both qualitative and quantitative factors in shaping how views are experienced, incorporating physiological indicators such as skin conductance, heart rate variability, and eye tracking to assess stress responses associated with various window configurations. Technological developments in tools like Electroencephalography (EEG) and fMRI have made it possible to objectively assess attention restoration by monitoring neurophysiological activity and neurochemical markers during exposure to nature [17].
Nevertheless, to date, no study has utilized EEG to directly examine emotional responses to classroom window views. This gap is significant, given the growing capabilities of EEG in recognizing, predicting, and classifying emotional states with precision [18]. Furthermore, advances in Virtual Reality (VR) technology make it possible to accurately simulate spatial conditions and visual environments, including various window views.
This study aims to objectively evaluate the cognitive and emotional responses elicited by different classroom window views - forest, park, and city - using EEG within an immersive VR setting. By combining VR simulation with EEG-based neurophysiological analysis, this research seeks to provide empirical insights into the restorative effects of natural visual exposure in classrooms, thereby contributing to the design of biophilic and cognitively supportive educational environments in green campus contexts.
2. Literature review
2.1. Classroom windows for energy efficiency and well-being
Classroom windows play a vital role in energy efficiency and student well-being by improving air quality, reducing energy consumption, and enhancing comfort, which supports learning outcomes [19,20]. While natural light improves visual comfort, it can also increase cooling demands [21], making a balance between daylight use and energy efficiency crucial, especially as students spend about 30% of their time in classrooms.
Windows can increase annual energy consumption by 6% to 181%, depending on factors such as orientation, glazing, and size [22]. In warm climates, efficient window design is particularly important, with key factors including window orientation, window-to-wall ratio, glazing type, and size [23]. Indonesia’s green building standard, for example, sets an Overall Thermal Transfer Value (OTTV) limit of 35 W/m² to optimize energy performance [24].
Adequate light levels, glare control, and outdoor views further enhance learning environments. Moreover, access to nature through windows improves pupil behaviour, well-being, and focus [8,10,11]. Additionally, natural views from windows are associated with better focus among students [9].
LEED v4.1 requires 75% of occupied areas to have outdoor views through glazing with at least 40% visible light transmittance [25]. This guideline also adopted by the GBCI [26]. High-quality views enhance satisfaction and reduce discomfort, with natural scenery preferred over urban settings [12].
2.2. Biophilic design as a medium for attention restoration
Attention restoration plays a critical role in mitigating negative emotional states such as burnout, anxiety, and mental fatigue. Natural environments, particularly those reflecting biophilic design principles, offer a psychological refuge that fosters calmness, safety, and well-being, which function in ways comparable to therapeutic interventions [27,28]. These restorative settings have been associated with reductions in physiological stress indicators such as salivary cortisol, heart rate, and blood pressure, while enhancing resilience and emotional balance [27,28].
Sustaining attentional capacity is essential for long-term cognitive performance, innovation, and motivation. Biophilic environments support this by eliciting soft fascination, a form of involuntary attention that promotes mental recovery without cognitive effort. This aligns with ART, which emphasizes the restorative effects of environments that provide fascination, a sense of being away, coherence (extent), and compatibility between user needs and environmental affordances [28].
Research in environmental psychology has demonstrated that natural and hybrid environments outperform urban settings in restoring attention and improving mood [28]. Technological advancements such as EEG and fMRI have enabled objective assessment of attention restoration by tracking neurophysiological responses and neurochemical shifts (e.g., dopamine, serotonin, adrenaline) during nature exposure [28].
Empirical findings further show that even passive exposure to nature, such as viewing greenery through windows or images, enhances attentional control. For example, students with natural views from their dormitories demonstrated significantly better attention performance than those without such exposure [29]. Direct interaction with real natural settings has also been linked to increased attentional capacity, reflective thinking, and overall well-being, surpassing the effects of simulated environments [30].
Design factors within indoor learning environments also contribute to cognitive recovery. A study on preschool facilities emphasized the importance of natural lighting, ventilation, and spatial organization for children's development and comfort, promoting eco-friendly design that supports holistic well-being [15]. Similarly, Felsten emphasized that environments offering mental detachment, coherence, fascination, and compatibility tend to be more restorative, characteristics often found in biophilic spaces [30].
Biophilic design has gained traction in educational, healthcare, and urban planning due to its restorative benefits [16,31]. For instance, a study at the University of Tennessee at Chattanooga used EEG and Stroop tests to evaluate the cognitive effects of short mental breaks across different campus environments. Students who spent time in natural outdoor settings achieved higher Stroop test scores and relaxation levels than those in indoor spaces without nature exposure. Physiological indicators such as Heart Rate Variability (HRV) and Theta-Beta Ratio (TBR), key markers of attention control and stress, also supported the positive impact of biophilic exposure on cognitive performance [17,30,32].
2.3. Electroencephalography (EEG) responses to environmental stimuli
EEG is widely employed to examine neurophysiological responses to environmental stimuli, especially those related to emotion, attention, and cognitive engagement. It provides high temporal resolution but is sensitive to artifacts from cardiac (ECG), ocular (EOG), and muscular (EMG) activity, requiring careful preprocessing to extract meaningful signals [18].
Recent advances have introduced portable, wireless EEG systems such as Emotiv’s EPOC X, which features 14 gel-free sensors and Bluetooth connectivity, making it suitable for field research in environmental neuroscience. Factors such as age, gender, and sociocultural background can affect EEG responses, with gender differences, shaped by both biology and social context, playing a notable role in data interpretation [18].
EEG studies comparing natural and urban environments reveal that natural settings enhance cognitive performance, attention, and meditative states [30] supporting theories that nature facilitates attentional restoration by reducing overstimulation. These effects are reflected in distinct neural signatures. Alpha waves across the cortex are linked to relaxation and inward-focused attention, while Beta and Gamma activity in the frontal cortex indicate focused attention. Frontal asymmetry also reflects emotional valence, with greater left frontal activation corresponding to positive emotions and approach motivation, and greater right activation associated with negative affect such as frustration, anxiety, or withdrawal. Arousal is typically marked by increased high-frequency activity in the posterior cortex, and meditative states may involve Theta activity in the frontal region.
The Theta/Beta Ratio (TBR), once linked to Attention Deficit Disorder and Attention-Deficit Hyperactivity Disorder [32], is now seen as an index of cognitive disengagement and mind-wandering. Elevated frontal TBR reflects resting states and has been applied in studies on learning capacity [17], and test anxiety [33]. In this study, TBR is used as a neural marker of the restorative potential of different window views.
2.4. Virtual Reality in environmental design research
The significance of classroom window views has been increasingly acknowledged across various studies. Wączyńska, Sokoł, and Martyniuk-Pęczek [34] compared objective and subjective assessments of classroom views, while others have examined the influence of natural views and daylight on thermal comfort, health, and energy efficiency [13]. In parallel, preferences regarding windows and daylighting have been explored in diverse settings, from kindergartens in Brazil to classrooms in the United States, emphasizing their impact on academic performance and user well-being [20]. Methodologies in these studies range from physical simulations and surveys to on-site measurements, yet many still rely on subjective evaluations, which are susceptible to bias. Additionally, conventional 2D visualization models often fail to replicate the complexity of real-world spatial perception or optimize window design strategies [34-36].
To address these limitations, VR has emerged as a robust tool, enabling immersive and repeatable experiments under controlled conditions. VR offers stereoscopic vision, realistic fields of view, and interactive environments, making it suitable for analysing visual perception with enhanced accuracy and ecological validity [36,37]. Unlike traditional surrogate methods such as photographs or videos, VR allows for more nuanced emotional and cognitive engagement. Studies have shown that higher realism in VR environments, through detailed graphics and naturalistic interactions, can elicit stronger affective responses and more valid environmental evaluations [37]. Despite its advantages, VR applications must consider participants’ expectations and perceptions, as these may influence their responses. Methodological rigor, including pre- and post-exposure assessments, is critical for improving the reliability of affective measurements within virtual environments. The sense of presence, which is essential in eliciting emotional engagement, relies not only on immersive hardware (e.g., Oculus Rift) but also on the consistency of sensory information and user interactivity [38].
VR’s utility extends to clinical, affective, and social neuroscience, where it enhances the realism of cognitive and emotional assessments. By enabling real-time interaction within ecologically valid scenarios, VR bridges the gap between laboratory settings and real-world functionality [39]. In training contexts, factors such as psychological and ergonomic fidelity may surpass mere graphical realism in predicting effective skill transfer. Research suggests that Augmented Reality (AR) and Mixed Reality (MR) also hold promise for future simulation environments, blending physical and virtual stimuli to optimize presence and task performance [40].
In environmental perception studies, VR-based lighting simulation tools, such as photometrically accurate daylit models, have been validated as reliable surrogates for real settings. They demonstrate strong congruence between subjective impressions of space (e.g., pleasantness, complexity, and view satisfaction) in both real and virtual environments [41]. Technological advancements in liquid crystal devices and optical systems further enhance the realism, image quality, and compactness of VR displays, opening new avenues for architectural and neuropsychological research [42].
The integration of VR into lighting performance simulation is exemplified by tools such as VRGlare, which allows for real-time glare analysis within immersive environments. This system enables intuitive three-dimensional visualization of visual discomfort, supplemented by multi-sensory feedback through haptic and audio channels. Such innovations significantly enhance the architect’s ability to interpret and mitigate glare in situ, offering practical applications in both design and neuroscientific studies [43].
3. Methods
The study was structured into five distinct stages. Stage I involved developing the classroom layout, including the plan dimensions, floor level, and window specifications with glazing types suitable for the Indonesian context. Stage II focused on designing external shading based on window orientations. Stage III constructed VR environments featuring various view variants. Stage IV measured psychological responses using EEG techniques, and Stage V compiled and analysed the collected data.
3.1. Experimental setting
The study was conducted in classrooms of Indonesian campus buildings located south of the equator (approximately 6°–8° S). Since most campus buildings in Indonesia are medium-rise (4–5 floors), a third-floor classroom was chosen to represent typical daylighting and energy performance conditions. The classrooms employed a double-loaded corridor layout, which is generally more space-efficient than single-loaded designs. A pedagogical classroom with a capacity of 30 students was selected. Based on the standard allocation of 25–30 ft² per student [44], its dimensions were set at 8 m × 10.5 m (84 m²).
These windows were oriented toward the north and south to ensure balanced daylight distribution and maintain thermal stability. The type of glazing and the window-to-wall area ratio were controlled variables in this study. The glazing used had a U-value of 5.7 W/m²·K and a Solar Heat Gain Coefficient (SHGC) of 0.93, corresponding to the properties of 8 mm clear single glazing commonly applied in most educational buildings in Indonesia. The model included windows along the exterior walls, with the lower edge positioned above desk height. Window dimensions were determined using the Overall Thermal Transfer Value (OTTV) method (Equation 1), with a regulatory threshold of 35 W/m² [24]. However, efforts were made to achieve OTTV values below 30 W/m². As north-facing windows typically receive greater solar radiation than those facing south, external shading devices were installed to achieve comparable OTTV values and balance solar heat gains across orientations. The energy efficiency levels, represented by OTTV values, were validated using simulation tools such as EnergyPlus and the Radiance plug-in in DesignBuilder. These tools have been widely validated for their accuracy in simulating daylight performance and window energy efficiency [45-48].
where OTTV is overall thermal transfer value on the building skin of a certain orientation (W/m2), α is solar absorptance, Uw is thermal transmittance of opaque wall (W/m2.K), WWR is window-to-wall area ratio of a certain orientation, TDEk is equivalent temperature difference (K), SF is solar factor (W/m2), SC is shading coefficient, Uf is thermal transmittance of fenestration (W/m2.K), and ΔT is design temperature difference between outdoor and indoor (K).
Six VR models were developed by combining two window orientations (left-facing and right-facing) with the three view types (forest, park, city) illustrated in Fig. 1 and Fig. 2. To ensure consistency with the daylighting conditions modeled in DesignBuilder, each VR environment was configured using identical geographic coordinates and sky model parameters. Specifically, an overcast sky condition with a fixed solar altitude angle of 0° was used. The tree species used in the forest and park views were selected from local varieties commonly found in natural forests surrounding the campus and in urban parks. For the forest view, the trees were arranged in a random configuration to reflect natural conditions. These simulations provided immersive and standardized visual environments for the EEG sessions.
Figure 1

Fig. 1. Classroom window views: (a) forest view on the left side, (b) forest view on the right side, (c) city view on the left side, (d) city view on the right side, (e) park view on the left side, (f) park view on the right side.
Figure 2

Fig. 2. Classroom furniture layout and seating position of the student/participant: (a) without shading, (b) with shading.
3.2. Participants and procedure
Ten architecture students (five male, five female), all aged 21 and without any visual impairments (Table 1), voluntarily participated in the study. The sample size aligns with previous neuroarchitecture research [49-52]. Participants were instructed to arrive with clean, unbound hair to ensure optimal EEG electrode contact. Each participant was fitted with a 14-channel Emotiv EPOC X EEG cap (Fig. 3), and conductive gel was applied to improve signal quality.
Table 1
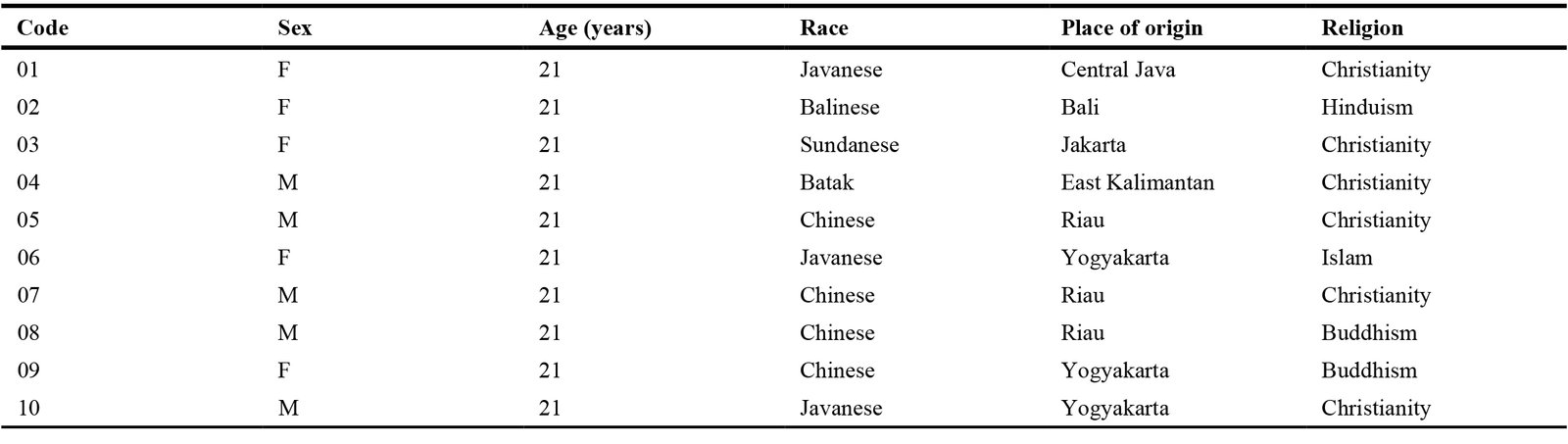
Table 1. Participants’ profiles.
Figure 3
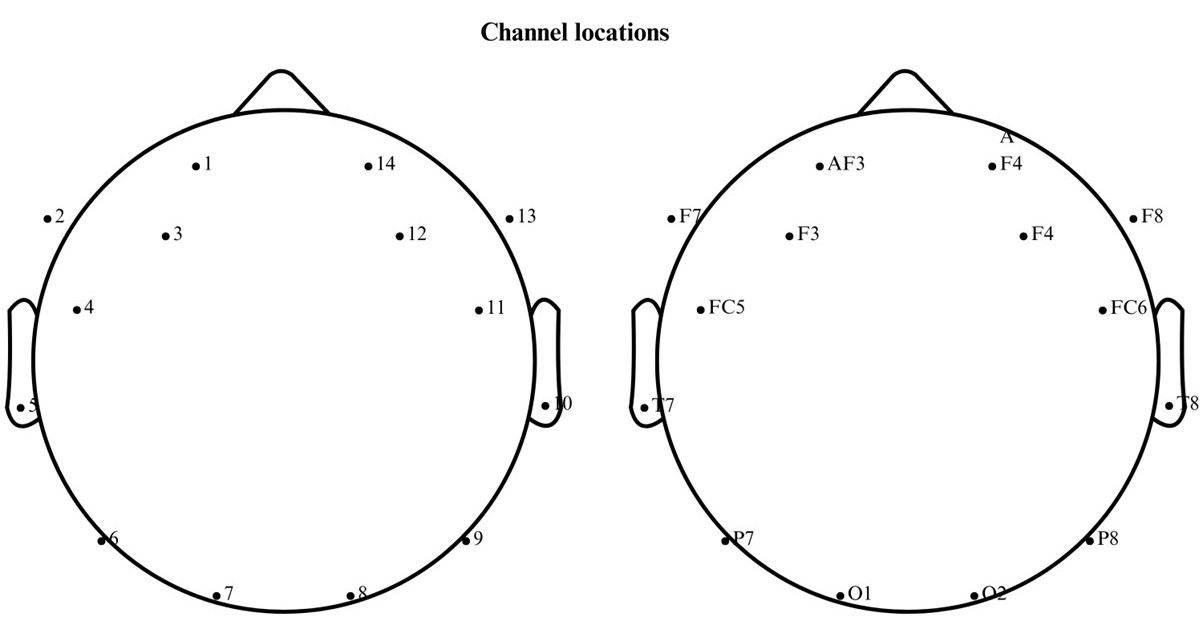
Fig. 3. The EEG channel location.
Participants completed six five-minute EEG sessions, each corresponding to one of the VR classroom models, with a five-minute rest period between changes in view type or window orientation. A five-minute duration is commonly used for EEG-based neural response measurements [53,54]. Prolonged exposure to intense VR stimuli may affect brainwave patterns, particularly in the frontal lobe [54]. All sessions were conducted in a controlled indoor environment to minimize external stimuli and distractions.
We identified five studies that explore the influence of cultural background on EEG-based emotional responses. Prior research shows that culture can shape neural emotion processing, as reflected in Late Positive Potential (LPP) differences between Dutch and Chinese participants [55], and between Asian and European American groups using Event-Related Potential methods [56]. Kraus et al. [57] found that Japanese participants with high interdependent self-construal exhibited reduced LPP when suppressing emotions, suggesting cultural values enhance regulatory effectiveness. Pugh et al. [58] further demonstrated that cultural and contextual factors influence fear processing. In contrast, [59] reported minimal cultural impact on EEG-based emotion recognition from audio stimuli.
Given the importance of cultural context in interpreting neural responses, Table 1 includes detailed participant data on ethnicity, place of origin, and religion. However, as all participants in this study share an Asian background, ethnic variability is unlikely to have significantly influenced their EEG responses to emotional stimuli.
3.3. EEG data collection and preprocessing
EEG signals were recorded across five standard frequency bands - Theta, Alpha, Low Beta, High Beta, and Gamma - yielding approximately 600 data points per one-minute segment. Raw EEG data from 14 scalp electrodes were exported in Excel format, then cleaned to remove formatting inconsistencies and structured for compatibility with Jamovi for statistical testing.
For each participant, data from both viewing orientations (left/right) and all view types were aggregated. EEG channels were grouped into three cortical regions - Frontal, Parietal, and Occipital - and averaged within each frequency band to produce 15 summary metrics (e.g., Frontal Alpha, Parietal Gamma). The resulting variables were coded to represent view type and window orientation.
3.4. Statistical analysis
Descriptive statistics were calculated, and data normality was assessed using the Shapiro-Wilk test. For data meeting normality criteria (p > 0.05), a Repeated Measures ANOVA (RM ANOVA) was conducted to evaluate within-subject effects of cortical region, frequency band, and window orientation. Post-hoc tests with Tukey correction and marginal means plots were generated to further explore significant interactions [60].
3.5. Advanced EEG processing in EEGLAB
In addition to the statistical analysis, EEG data underwent a rigorous 12-step preprocessing procedure using the EEGLAB toolbox in MATLAB to ensure signal clarity and reliability. The workflow began with raw data import and application of a band-pass filter (1-40 Hz) to remove low-frequency drifts and high-frequency noise while preserving neural activity relevant to cognitive and emotional processing. Subsequent steps included down-sampling, line noise removal, bad channel detection, re-referencing, and segmentation into epochs suitable for analysis.
Artifact removal was achieved through a two-stage application of Independent Component Analysis (ICA). The first ICA identified and eliminated common artifacts such as eye blinks and muscle movements, followed by the rejection of noisy epochs. A second ICA was then performed to refine the decomposition further. Finally, source localization was conducted using equivalent current dipole fitting, and the cleaned data were saved for subsequent analysis. This comprehensive pipeline effectively removed non-neural noise, enhancing the interpretability of EEG signals across the different classroom view conditions.
4. Results
4.1. Energy and daylighting performance of each window model
Using Equation 1, the building envelope of a campus classroom with a south-facing (right-side) window measuring 10.5 m × 2.1 m and positioned 1 m above the floor in an 84 m² classroom has an OTTV of 22.06 W/m². This OTTV value is considered optimal, qualifying the building as a green building. The same window dimensions, height, and materials were applied to a classroom with a north-facing window. To maintain a comparable OTTV, a 1m-wide shading device was installed above the north-facing window. As a result, the OTTV for the north-facing window was 21.07 W/m², which is 4.4% lower.
Energy performance simulations of both classroom models using EnergyPlus revealed a relatively small difference in cooling load, with a reduction of 14.4%. The classroom with a right-side window recorded a cooling load of 18.84 kWh, while the classroom with a left-side window showed a lower load of 16.13 kWh. Daylighting performance was evaluated using Radiance simulations in DesignBuilder. Although both classrooms exhibited similar illuminance levels, the south-facing window provided a more uniform distribution of daylight. The right-side window produced a daylight factor ranging from 0% to 27% and horizontal illuminance levels between 0 lux and 2701 lux. In contrast, the left-side window yielded a daylight factor between 0% and 17%, with illuminance levels ranging from 0 lux to 1728 lux. Annual daylighting performance, assessed through daylight autonomy (DA), was nearly identical for both window orientations, ranging from 80% to 90% (Fig. 4).
Figure 4
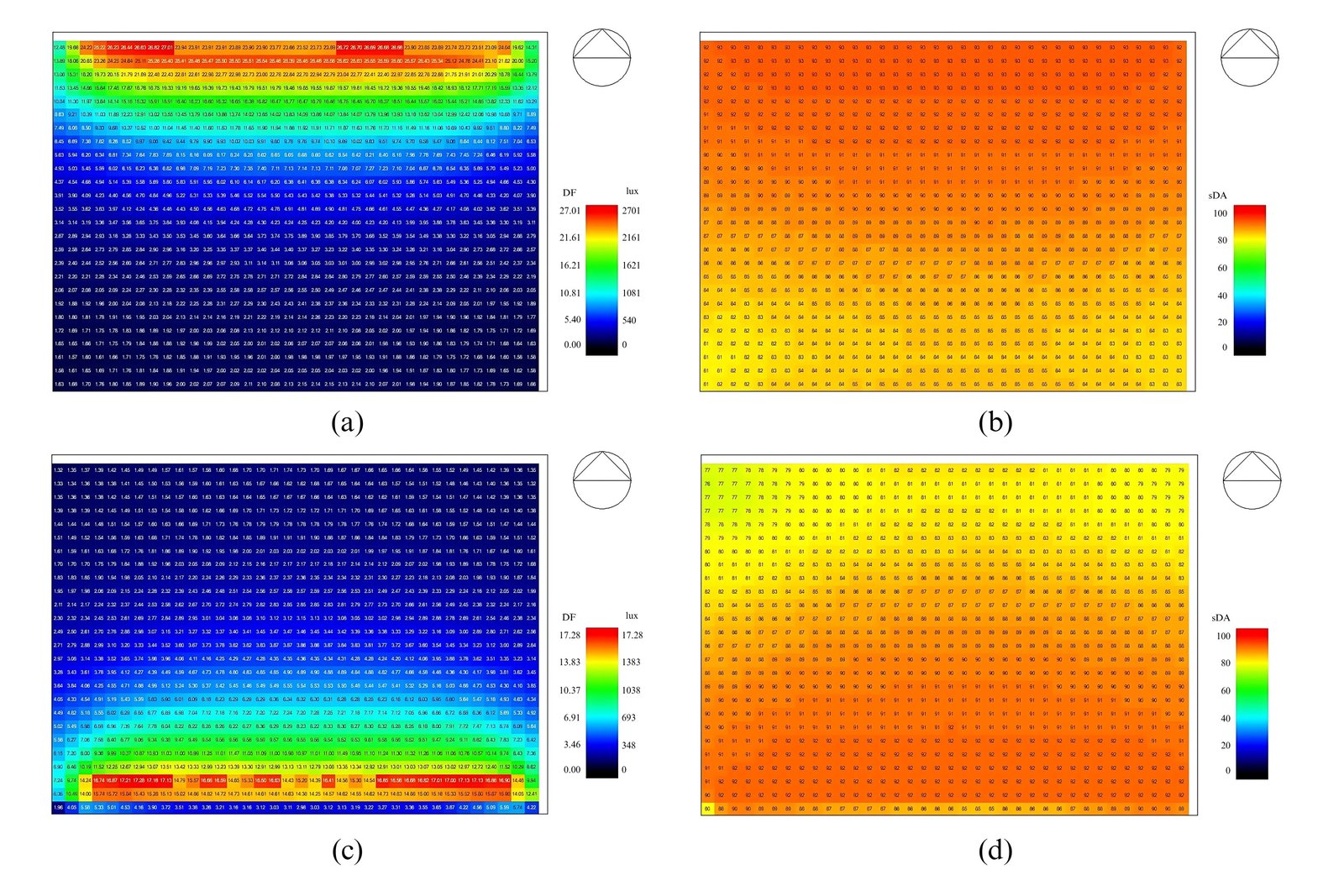
Fig. 4. (a) Simulation of daylight factor and illuminance of a classroom with right-side window, (b) Simulation of daylight autonomy of a classroom with right-side window, (c) Simulation of daylight factor and illuminance of a classroom with left-side window, (d) Simulation of daylight autonomy of a classroom with left-side window.
4.2. Results of the normal distribution tests
Before conducting the marginal means estimation analysis, a normality test was performed for each EEG measurement to ensure that the data or variables were normally distributed. Table 2 presents the results of the normality test for all participants, window orientations, and view types, indicating that all variables in all measurements were normally distributed (Shapiro-Wilk p < 0.05). Therefore, the research proceeded to the next stage of analysis.
Table 2
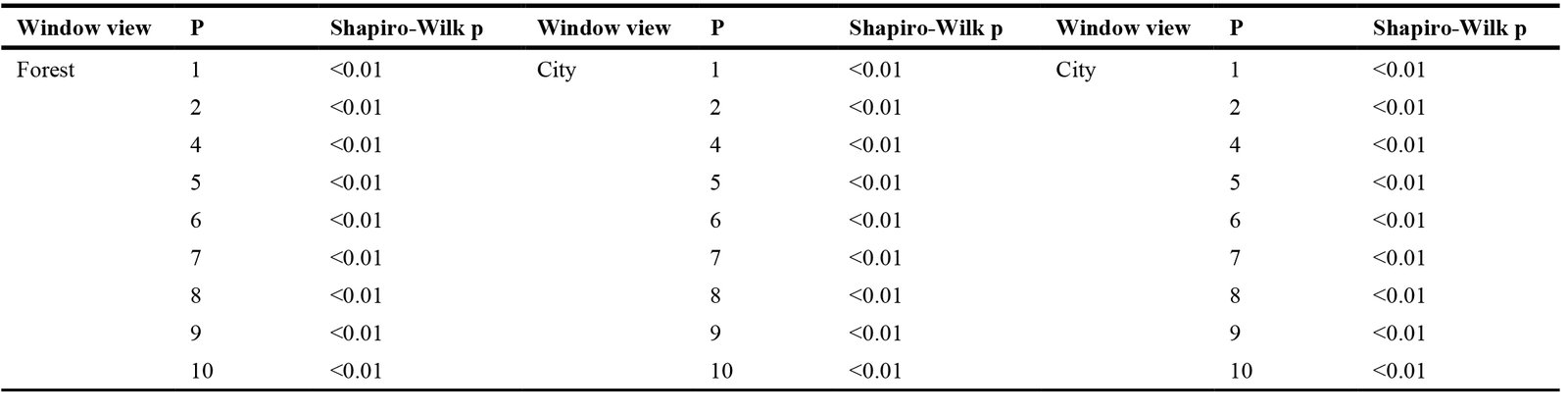
Table 2. Results of normal distribution tests.
4.3. Results of repeated measure ANOVA
The results of the repeated ANOVA analyses revealed consistent neurophysiological patterns across participants when exposed to different classroom window views. For Participant 01, all three visual conditions - forest, city, and park - elicited significant differences in brain activity, particularly in the frontal, right temporal, and occipital regions (F > 127; p < 0.001). EEG responses also varied significantly across wavebands (Theta to Gamma), and the window position (left vs. right) demonstrated notable influence (F = 32-98; p < 0.001), indicating that certain spatial orientations may enhance neural processing.
Similarly, Participant 02 exhibited significant neural variation across all variables for each view. Brain regions showed F-values ranging from 127 to 302, wavebands from 67 to 154, and window position from 31 to 96 (all p < 0.001), reflecting a strong modulation of brain dynamics depending on the visual stimulus and spatial exposure. A comparable pattern was found in Participant 03, whose responses were consistently significant with brain region F-values ranging from 168 to 791, EEG wavebands from 240 to 662, and window orientation effects ranging from 88 to 166 (p < 0.001), again emphasizing the influence of view type and direction.
The response from Participant 04 was notably pronounced, especially under the forest view, which yielded extremely high F-values - 6395 for brain region, 1854 for waveband, and 852 for window position - demonstrating an exceptional sensitivity to natural scenes. City and park views produced similarly significant, albeit lower, values. In the case of Participant 05, strong effects were also observed, particularly under the park view, where brain activity and EEG waveband responses peaked (F = 1063 and F = 407, respectively), suggesting heightened cognitive and perceptual engagement with green environments.
Participant 06 showed consistent and significant results across all viewing conditions, with the forest view generating the highest F-values for both brain activity (F = 1679) and EEG wavebands (F = 765), while the influence of window position remained evident. For Participant 07, although F-values were generally lower (brain region F = 36-86; wavebands F = 108-401), all differences remained statistically significant, indicating that even moderate shifts in environmental stimuli and spatial configuration can affect neural responses.
In the case of Participant 08, brain activation (F = 266-765), EEG waveband responses (F = 74-521), and window orientation (F = 56-111) were significantly affected by all three types of views, with forest and park conditions showing a more restorative effect compared to urban scenes. Participant 09 responded more strongly to forest and park views, reflected in high F-values for brain regions (F = 323-1105) and EEG wavebands (F = 362-708), while the effect of window position was significant but varied in magnitude (F = 13-271).
Lastly, Participant 10 demonstrated the most prominent responses under the park view, where brain activity (F = 330), EEG waveband dynamics (F = 1588), and window position (F = 613) reached peak significance. Forest and city views also produced meaningful changes, underscoring the consistent influence of environmental view type and spatial orientation on EEG-based brain activity.
4.4. The EEG spectral and topographic maps
The results of EEG data preprocessing were presented through spectral graphs and topographic brain maps for each participant, corresponding to different classroom settings based on the window side and view. Power spectral density was visualized as a graph plotted against frequency, accompanied by five representative topographic maps selected from the Delta (1-3 Hz), Theta (4-8 Hz), Alpha (8-13 Hz), Beta (13-30 Hz), and Gamma (30-50 Hz) frequency bands.
Figure 5 shows that the electrophysiological data revealed that the forest view elicited the highest Alpha peak amplitude in Participant 1 (M = 15.8 µV²/Hz, SD = 4.1), significantly surpassing the city view (M = 10.2, SD = 3.5; t(n-1) = 3.97, p < .01), and was accompanied by robust Alpha (8-13 Hz) and Theta (4-8 Hz) activity across parietal and occipital regions - patterns closely associated with relaxed alertness, internalized attention, and restorative cognitive states. Topographical maps indicated symmetrical high-frequency synchronization over posterior areas, supporting visually restful processing. Beta band (13-30 Hz) activity was also significantly higher frontally in the forest condition compared to the city (t(n-1) = 2.84, p = 0.03), while the city view showed attenuated Alpha and increased Gamma (>30 Hz) activity over frontal-temporal areas, suggesting cognitive overstimulation and reduced sensory restoration due to complex urban stimuli. The park condition, with intermediate Alpha (M = 13.2 µV²/Hz, SD = 3.2) and Beta patterns, showed no significant difference from the forest (p = 0.12) but was significantly higher than the city (p < 0.05), indicating partial restorative benefits and a balanced state of relaxed yet alert engagement. When viewing the right-side window, Participant 1 again exhibited the most restorative EEG profile during the forest view. This was characterized by enhanced low-frequency (Delta/Theta) power and strong posterior Alpha activation, consistent with visual rest and reduced cognitive demand. The city views triggered frontal Beta and Gamma activity, while the park view revealed a balanced neural state, combining mild relaxation and active attention.
Figure 5
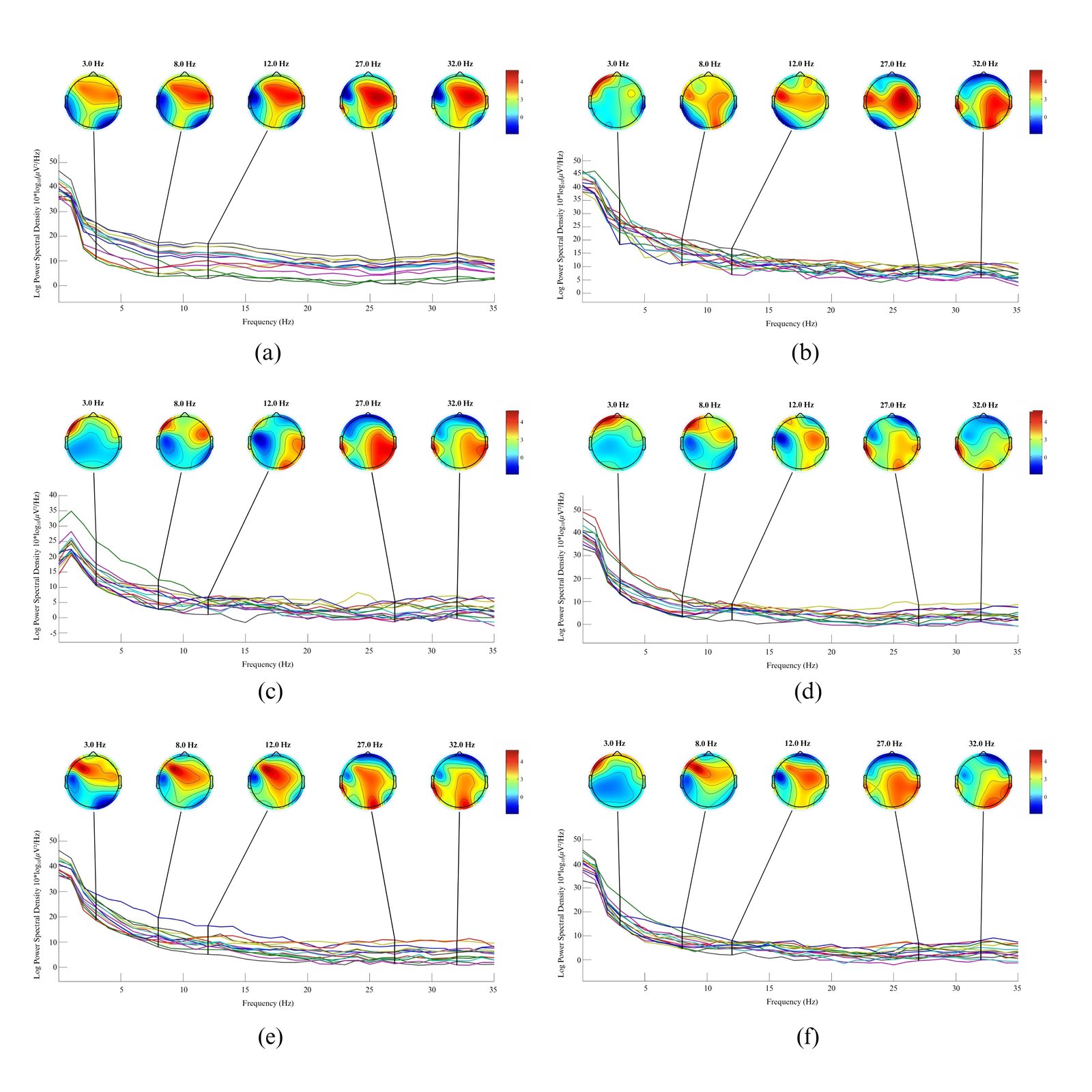
Fig. 5. Topographical maps of Participant 1’s brain responses to left-side windows with forest (a), city (c), and park (e) views, and right-side windows with forest (b), city (d), and park (f) views.
In Participant 2, the forest-view condition showed the highest overall power spectral density across all frequency bands, particularly in the low (3-8 Hz) and mid-frequency ranges (12 Hz), with strong activation in occipital and parietal regions suggesting enhanced visual processing and attentional states. The city-view condition showed a marked reduction in EEG power, especially in lower frequencies, alongside a more dispersed and less intense activation pattern, potentially reflecting increased cognitive load. The park-view condition exhibited intermediate spectral power with balanced frontal and occipital activation. With a right-side window, the forest view fostered posterior Alpha-Theta dominance and minimal Beta/Gamma activity. In contrast, the city views increased frontal Beta and Gamma power and suppressed posterior Alpha. The park view exhibited moderate posterior Alpha and Theta with neutral Beta/Gamma patterns, representing soft fascination and sustained attention without fatigue.
Participant 3 showed increased posterior Alpha (8-12 Hz) and Gamma (32 Hz) under the forest view, indicating a calm and restorative attentional state. The city view induced lower Alpha power and enhanced frontal Gamma, indicative of heightened cognitive load. The park view reflected a middle ground with moderate Alpha and balanced Gamma across bilateral regions (Fig. 6). With a right-side window, the forest condition maintained posterior Delta-Theta activity and occipital-parietal Alpha, while city-view exposure suppressed Alpha and elevated frontal high-frequency bands. The park view showed posterior-dominant low-frequency activity and moderate Gamma, suggesting relaxation with attentional engagement.
Figure 6
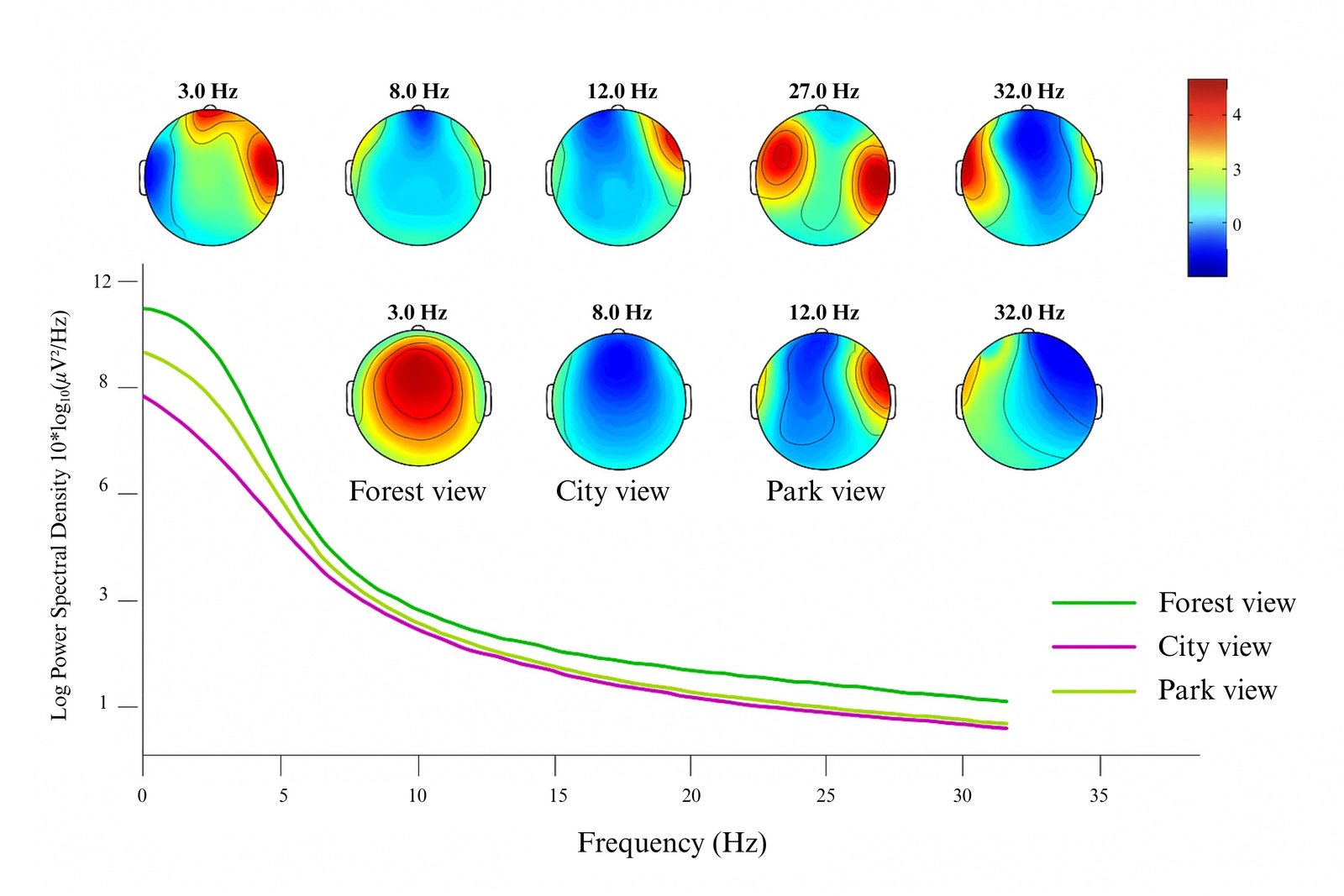
Fig. 6. Merged topographical maps of Participant 3’s brain responses to left-side windows with forest, city, and park views.
In Participant 4, forest-view exposure elicited relatively stable activity in Alpha and Beta bands, suggesting a relaxed yet alert mental state. The city view showed elevated Beta and low Gamma power, pointing to increased arousal and cognitive load. The park view produced intermediate spectral features, balancing arousal and engagement. With a right-side window, the forest view led to robust posterior Alpha-Theta and minimal frontal high-frequency activation. The city view induced elevated Beta-Gamma in frontal-temporal areas, while the park condition displayed moderate Alpha-Theta and mild high-frequency responses.
Participant 5’s response to the forest view was characterized by mild posterior Delta activity, pronounced Alpha, and lateralized sensorimotor alpha rhythm (mu rhythm). Low Beta and localized Gamma activity further reinforced an overall restorative pattern. In contrast, the city view elicited posterior Delta suggestive of fatigue, suppressed frontal Alpha, occipital–temporal sensorimotor alpha overload, and elevated Beta/Gamma indicative of overstimulation. The park view showed central Delta suppression, balanced Alpha and sensorimotor alpha activity, mild Beta, and modest Gamma (Fig. 7). On the right side, the forest view generated a relaxed attentional state with moderate low-frequency power and minimal high-frequency activity. The city view elevated frontal-temporal Beta/Gamma with right-lateralized Alpha, while the park view showed balanced Theta/Alpha and low Gamma, consistent with softly engaging environments.
Figure 7
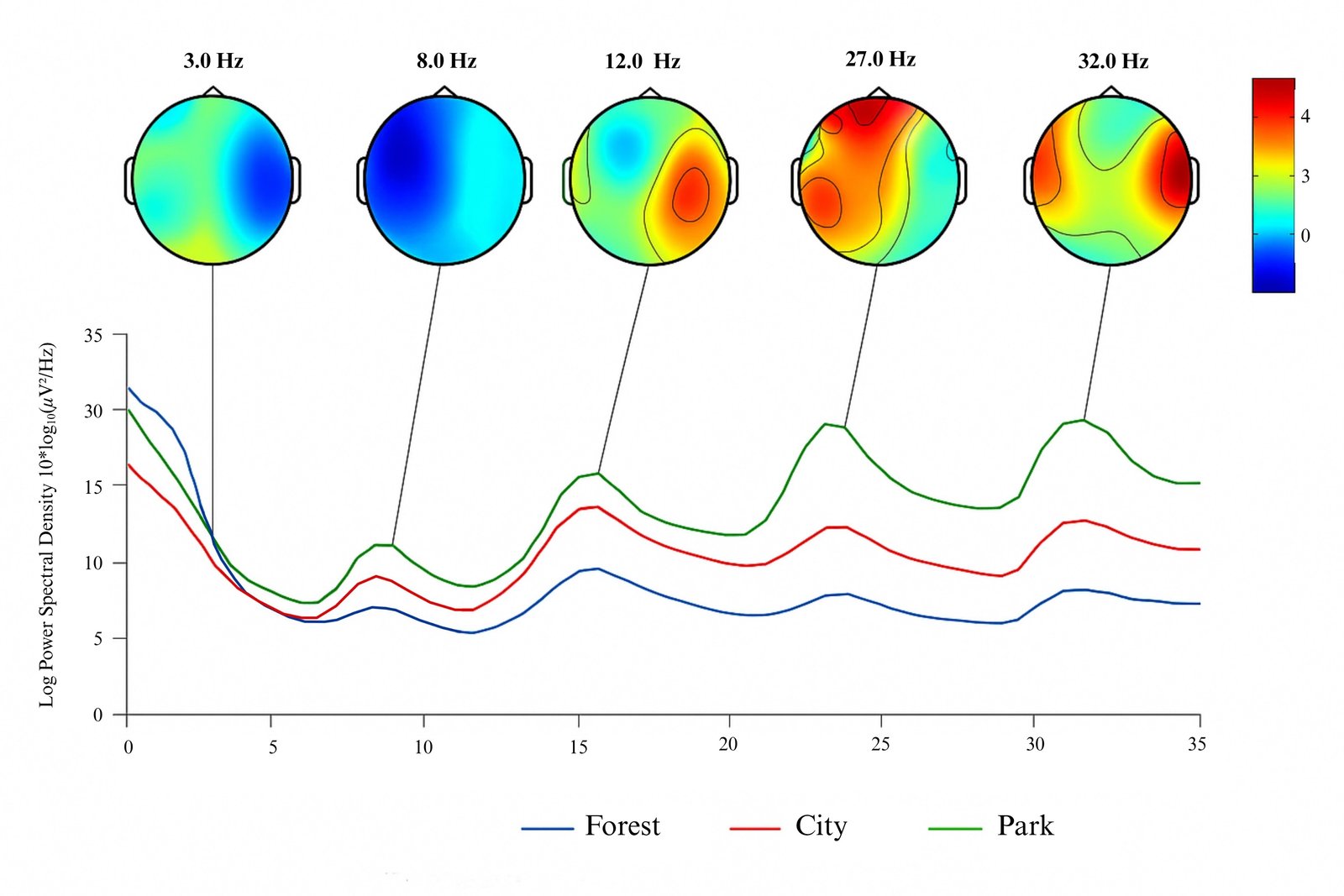
Fig. 7. Merged topographical maps of Participant 5’s brain responses to left-side windows with forest, city, and park views.
Participant 6 displayed strong posterior Alpha and moderate low-frequency power under the forest view, with balanced Beta/Gamma indicating non-stressful attentiveness. The city view showed suppressed Delta/Theta and Alpha, and increased frontal Beta/Gamma with left asymmetry, pointing to cognitive overload. The park view showed high Delta/Theta, moderate Alpha with frontal spread, and mild Beta/Gamma, with symmetrical topographies. With a right-side window, forest views produced higher low-frequency power (Delta/Theta) and moderate posterior Alpha. The city view elevated frontal Beta/Gamma with reduced low-frequency activity, while the park view showed modest Alpha/Theta and balanced topographies.
In Participant 7, forest-view conditions elevated low-frequency bands and posterior activity, indicating a relaxed, restorative state. City views induced higher Beta/Gamma in frontal-temporal regions, suggesting arousal. Park views displayed moderate power across bands and balanced activation, implying relaxed alertness (Fig. 8). On the right side, forest views increased Delta/Theta with bilateral posterior Alpha peaks. City views showed elevated Beta/Gamma and reduced Alpha. Park views showed moderate activation and attentional calm. These patterns affirm that natural views - especially dense greenery - foster restorative brain states through enhanced posterior low-frequency oscillations, while urban views heighten cognitive demands.
Figure 8
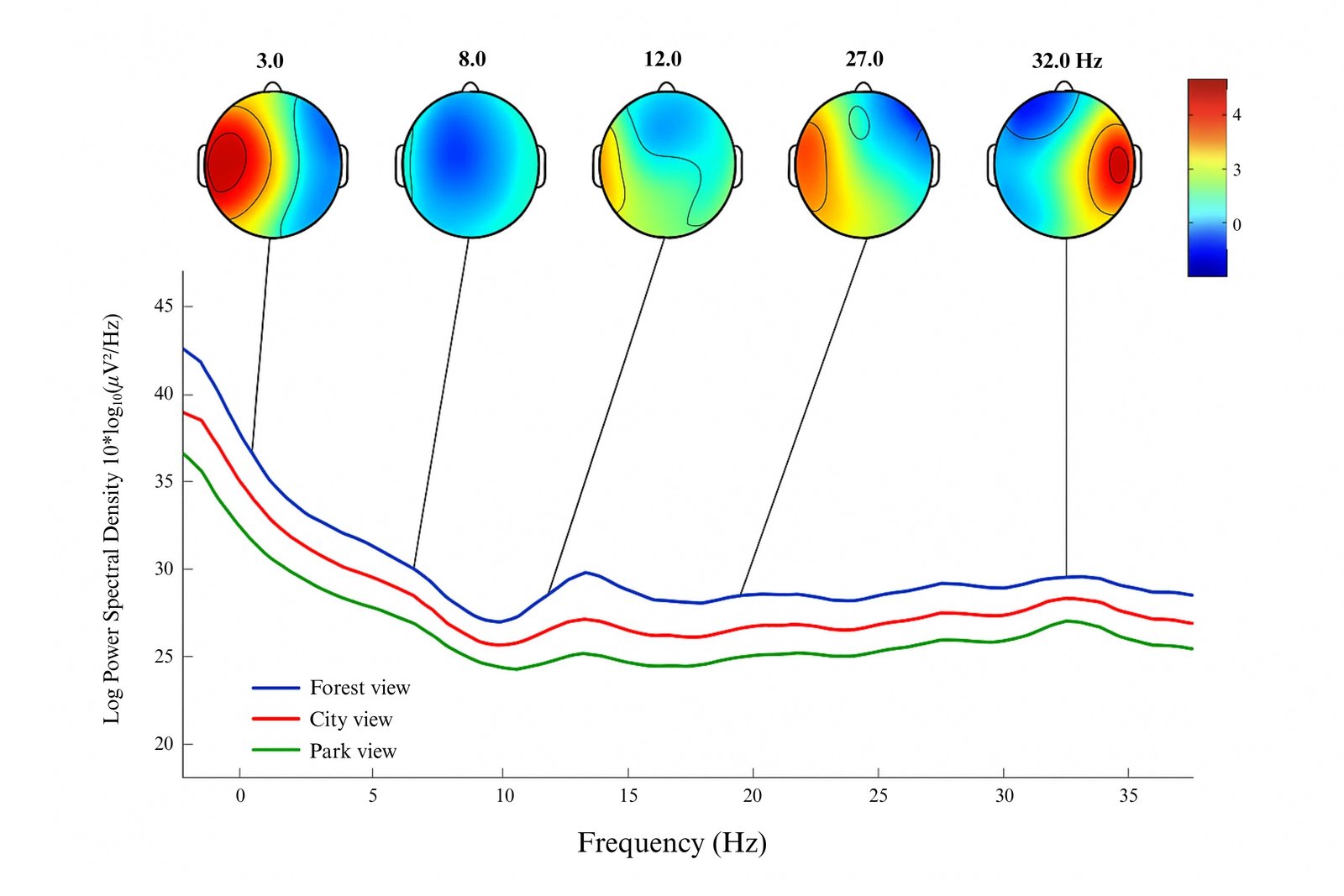
Fig. 8. Merged topographical maps of Participant 7’s brain responses to left-side windows with forest, city, and park views.
For Participant 8, left-side forest-view exposure showed high low-frequency PSD values (3 Hz, 8 Hz) over frontal and occipital regions, suggesting a relaxed, attentive state. City views showed reduced low-frequency and increased high-frequency activity, indicative of overstimulation. Park views had balanced Theta, Alpha, and Beta, with occipital and frontal activation. On the right, forest views again evoked strong Delta/Theta power with posterior dominance. City views produced high Beta/Gamma over frontal-lateral areas. The park condition demonstrated moderate spectral power and symmetrical topographies.
Participant 9 showed strong posterior Delta and Alpha in forest-view conditions, consistent with visual rest. City views induced frontal-temporal Beta/Gamma and Alpha suppression, indicating high cognitive load. Park views presented moderate low-frequency activity and distributed Alpha. On the right side, forest views showed suppressed spectral power and strong posterior calm. City views elicited high Beta and Gamma activity with widespread cortical activation. In contrast, park views showed moderate Alpha and frontal Beta activity, suggesting a restorative yet cognitively engaging profile.
In Participant 10, forest views yielded bilateral occipital Delta and posterior Alpha, indicating restorative processing. City views caused central suppression, frontal-parietal Theta, and increased Beta/Gamma, consistent with stimulation. The park view showed Delta calm, posterior Alpha, and moderate Beta/Gamma activation. On the right side, the forest view elicited high PSD across all bands, with right parietal and occipital activation indicative of relaxed alertness. City views were more diffuse with frontal Alpha suppression and mild Beta activity, while the park view displayed balanced Alpha-Beta with posterior emphasis and lower Gamma, suggesting partial restoration.
Across participants, the EEG data consistently indicate that forest views promote restorative neural states characterized by Alpha-Theta enhancement, posterior activation, and reduced high-frequency load. City views, conversely, produce overstimulation, reflected in elevated frontal Beta-Gamma and suppressed Alpha. Park views emerge as a cognitive middle ground, supporting both attentional engagement and neural recovery. These findings highlight the importance of integrating natural elements, particularly forest-like greenery, into learning environments to promote cognitive restoration. This aligns with core principles of neuroarchitecture and biophilic design.
5. Discussion
The findings of this study on the neurophysiological impact of different classroom window views - forest, park, and city - within energy-efficient, VR-simulated environments, as measured by EEG, reveal consistent patterns across participants that underscore the restorative value of nature-based visual stimuli. These results support key theoretical frameworks, particularly ART and principles of biophilic design [61–63].
Forest views elicited the most favorable EEG responses, marked by increased Alpha and Theta activity in the posterior brain regions, reduced high-frequency (Beta/Gamma) activation in the frontal areas, and more symmetrical topographical patterns. These neural signatures are typically linked to relaxed alertness, internalized attention, and visual rest, reinforcing the forest view as the most restorative environment. The results align with previous research showing that dense greenery promotes mental restoration and attentional recovery through soft, passive fascination, a key feature of biophilic environments [61–64].
In contrast, city views induced heightened Beta and Gamma activity in the frontal-temporal areas, suppressed Alpha rhythms, and asymmetric brain activation, particularly in the left hemisphere. These responses suggest increased cognitive load and environmental overstimulation resulting from the visual complexity and unpredictability of urban settings. Such neural patterns reflect a shift toward externally oriented attention and task-driven mental states, which may impair sustained attention and emotional regulation in classroom settings.
The park view consistently yielded EEG responses that were intermediate between the forest and city conditions. Moderate Alpha and Theta activity, balanced Beta power, and broadly distributed brain activation suggest a state of mindful engagement and partial restoration. These findings align with the concept of "soft fascination," whereby semi-natural environments provide enough visual interest to engage attention without overwhelming the sensory system, thus supporting cognitive performance and psychological well-being [64].
Regarding window orientation (left vs. right side), no significant differences were observed in the restorative effects of the views, although minor variations in hemispheric activation were detected in some cases. This suggests that the type and quality of the external view are more influential than window orientation in shaping psychological responses. This is especially true when lighting conditions are equalized through shading and passive thermal strategies, as applied in this study. However, when window position leads to differences in daylight quantity and quality, it may significantly affect EEG patterns.
Overall, the results highlight the potential for natural views, especially forest landscapes, to function as cognitive and emotional regulators in learning environments. By incorporating green elements into students’ visual fields through strategic window design, educational institutions may enhance attention, reduce mental fatigue, and support overall psychological well-being. These findings also contribute valuable insight to the design prescriptions for window views in green educational buildings.
Additionally, the use of VR environments and EEG measurements in this study demonstrates the value of combining immersive simulations with objective physiological metrics to investigate environmental psychology in architectural contexts. However, the study did not account for several factors critical to successful immersion, including inclusivity, extensiveness, surrounding scope, and vividness. Nor did it explore the roles of immersive tendency, personal relevance of the VR content, or the type of VR display used. Consistent with previous findings [65], immersive tendency did not significantly affect the outcomes in this study.
Although participants represented diverse ethnic, regional, and religious backgrounds, their EEG responses to the three view types were broadly consistent. This indicates that cultural background did not substantially shape neurophysiological reactions to visual environmental stimuli. The shared educational context, similar age group, and inherently restorative quality of natural views likely reduced cultural variation in responses. Thus, the type and quality of visual exposure appeared more influential than cultural identity in determining EEG patterns.
Nonetheless, several limitations should be acknowledged. The relatively small sample size, while comparable to earlier neuroarchitecture research, constrains the generalizability of the findings. Furthermore, individual differences - such as prior experience with natural environments, current stress levels, and personality traits - may mediate EEG responses and deserve further exploration.
Future research should include larger and more diverse participant groups, and assessments in real-world classrooms with varied window configurations and external views. Advancing the use of mobile EEG technologies and improving the realism of VR simulations can help bridge the gap between controlled laboratory studies and real-world architectural application. Integrating physiological data with behavioural measures and academic outcomes will also provide a more comprehensive understanding of how biophilic classroom design influences learning and well-being.
6. Conclusion
This study provides empirical evidence that the quality of window views in classrooms significantly affects students’ neurophysiological responses, particularly in green campus environments that meet energy-efficient design standards. Using VR simulations and EEG measurements, forest views were found to consistently elicit restorative brain activity, marked by elevated Alpha and Theta power and reduced Beta and Gamma frequencies. These patterns are associated with relaxed alertness and cognitive restoration, supporting principles from Attention Restoration Theory and biophilic design.
In contrast, city views triggered neural markers of overstimulation and cognitive load, including increased frontal Beta and Gamma activity and suppressed Alpha rhythms. Park views produced intermediate effects, indicating partial restoration and attentional engagement, supporting the concept of “soft fascination.” The influence of window orientation was minimal when lighting and thermal conditions were equalized, underscoring that view content and quality are more critical than spatial placement in determining psychological outcomes.
These findings highlight the importance of incorporating greenery-rich, natural views into classroom design to support student focus, reduce mental fatigue, and enhance overall well-being. Moreover, they demonstrate the value of combining immersive VR environments with objective neurophysiological tools like EEG to evaluate design interventions in educational architecture.
Importantly, this evidence has direct implications for educational policy and architectural practice. Educational authorities and school planners should consider visual access to nature as a fundamental aspect of student health and cognitive development, rather than a secondary design feature. Window placement and outdoor landscape planning should be strategically aligned to maximize exposure to restorative natural elements, especially forest-like greenery. These insights can inform evidence-based architectural guidelines for the design of biophilic, student-centered, and sustainable learning environments.
To build upon these findings, future research should involve larger and more diverse participant groups, and incorporate behavioural and academic performance metrics. The integration of physiological, psychological, and educational data will further support the development of comprehensive policies and green building standards that promote cognitive restoration, attentional resilience, and emotional well-being in academic settings.
Acknowledgment
The authors gratefully acknowledge the research funding provided by the Research and Community Service Centre, Universitas Atma Jaya Yogyakarta (85/LPPM-Pen/In). The authors also made use of an AI-based language tool (ChatGPT) to assist with language editing and polishing of this manuscript.
Funding
This research received no external funding.
Contributions
Floriberta Binarti contributed to Conceptualization, Methodology, Formal analysis, Investigation, Resources, Writing– review and editing, and Supervision. Nimas Sekarlangit contributed to Methodology, Investigation, Formal analysis, Data curation. Meita Kasianus Virgin Brilianto contributed to Investigation, Software, Formal analysis, Data curation. Bhanu Rizfa Hakim contributed to Visualization.
Declaration of competing interest
The authors declare no conflict of interest.
References
- H. Han, K. Kiatkawsin, W. Kim, J.H. Hong, Physical classroom environment and student satisfaction with courses, Assessment & Evaluation in Higher Education, 43:1 (2018), 110-125. https://doi.org/10.1080/02602938.2017.1299855
- S. Jin, L. Peng, Classroom perception in higher education: The impact of spatial factors on student satisfaction in lecture versus active learning classrooms, Frontiers in Psychology, 13 (2022). https://doi.org/10.3389/fpsyg.2022.941285
- N. van den Bogerd, S.C. Dijkstra, K. Tanja-Dijkstra, M.R. de Boer, J.C. Seidell, S.L. Koole, J. Maas, Greening the classroom: Three field experiments on the effects of indoor nature on students' attention, well-being, and perceived environmental quality, Building and Environment, 171 (2020) 106675. https://doi.org/10.1016/j.buildenv.2020.106675
- N. Castilla, C. Llinares, J.M. Bravo, V. Blanca, Subjective assessment of university classroom environment, Building and Environment, 122 (2017) 72-81. https://doi.org/10.1016/j.buildenv.2017.06.004
- Z. Yang, B. Becerik-Gerber, L. Mino, A study on student perceptions of higher education classrooms: Impact of classroom attributes on student satisfaction and performance, Building and Environment, 70 (2013) 171-188. https://doi.org/10.1016/j.buildenv.2013.08.030
- B.J. Al-Khatatbeh, S.N. Ma'Bdeh, Improving visual comfort and energy efficiency in existing classrooms using passive daylighting techniques, Energy Procedia, 136 (2017), 102-108. https://doi.org/10.1016/j.egypro.2017.10.294
- European Standard, Issue 93 Daylighting-European Standard EN 17037, Issue 93:October (2019), 17037 [Online]. Available: https://orf.od.nih.gov/TechnicalResources/Documents/Technical Bulletins/19TB/Daylighting - European Standard EN 17037 October 2019-Technical Bulletin_508.pdf [accessed: 01-Feb-2024].
- J. Kim, M. Kent, K. Kral, T. Dogan, Seemo: A new tool for early design window view satisfaction evaluation in residential buildings, Building and Environment, 214 (2022) 108909. https://doi.org/10.1016/j.buildenv.2022.108909
- P. Lindemann-Matthies, D. Benkowitz, F. Hellinger, Associations between the naturalness of window and interior classroom views, subjective well-being of primary school children and their performance in an attention and concentration test, Landscape and Urban Planning, 214 (2021) 104146. https://doi.org/10.1016/j.landurbplan.2021.104146
- B.S. Matusiak, No-greenery line and greenery-view factor, new architectural design tools, Journal of Daylighting, 7:2 (2020) 282-286. https://doi.org/10.15627/jd.2020.24
- A.L. Pearson, C.D. Brown, A. Reuben, N. Nicholls, K.A. Pfeiffer, K.A. Clevenger, Elementary Classroom Views of Nature Are Associated with Lower Child Externalizing Behavior Problems, International Journal of Environmental Research and Public Health, 20:9 (2023). https://doi.org/10.3390/ijerph20095653
- F. Abd-Alhamid, M. Kent, Y. Wu, Quantifying window view quality: A review on view perception assessment and representation methods, Building and Environment, 227:P2 (2023), 109742. https://doi.org/10.1016/j.buildenv.2022.109742
- Y. Jiang, N. Li, A. Yongga, W. Yan, Short-term effects of natural view and daylight from windows on thermal perception, health, and energy-saving potential, Building and Environment, 208:October (2022), 108575. https://doi.org/10.1016/j.buildenv.2021.108575
- H. Medhat Assem, L. Mohamed Khodeir, F. Fathy, Designing for human well-being: The integration of neuroarchitecture in design - A systematic review, Ain Shams Engineering Journal, 14:6 (2023), 102102. https://doi.org/10.1016/j.asej.2022.102102
- R. Omar, D.P. Rana, A.S. Rastogi, Enhancing Learning with Nature-Inspired Design in Schools, Town Planning and Management, 10:2 (2024), 18-27.
- R. Berto, Exposure to restorative environments helps restore attentional capacity, Journal of Environmental Psychology, 25:3 (2005), 249-259. https://doi.org/10.1016/j.jenvp.2005.07.001
- E.K. Swart, T.M.J. Nielen, S. Shaul, M.T. Sikkema-De Jong, Frontal Theta/beta-ratio (Tbr) as potential biomarker for attentional control during reading in healthy females, Cognition Brain, Behavior. An Interdisciplinary Journal, 24:3 (2020), 187-211. https://doi.org/10.24193/cbb.2020.24.11
- D. Dadebayev, W.W. Goh, E.X. Tan, EEG-based emotion recognition: Review of commercial EEG devices and machine learning techniques, Journal of King Saud University - Computer and Information Sciences, 34:7 (2022), 4385-4401. https://doi.org/10.1016/j.jksuci.2021.03.009
- N.G. Vasquez et al., Ventilation strategies and children's perception of the indoor environment in Swedish primary school classrooms, Building and Environment, 240 (2023). https://doi.org/10.1016/j.buildenv.2023.110450
- N.G. Vásquez, M.L. Felippe, F.O.R. Pereira, A. Kuhnen, Luminous and visual preferences of young children in their classrooms: Curtain use, artificial lighting and window views, Building and Environment, 152 (2019) 59-73. https://doi.org/10.1016/j.buildenv.2019.01.049
- A. Aniebietabasi, M. Donn, G. Thomas, The Influence of Indoor Environmental Quality in Schools, 51st International Conference of the Architectural Science Association, Architectural Science Association (ANZAScA), Wellington, New Zealand, February 2018, pp. 625-634.
- S.K. Alghoul, H.G. Rijabo, M.E. Mashena, Energy consumption in buildings: A correlation for the influence of window-to-wall ratio and window orientation in Tripoli, Libya, Journal of Building Engineering, 11 (2017) 82-86. https://doi.org/10.1016/j.jobe.2017.04.003
- R.L. Hwang, W.A. Chen, Identifying relative importance of solar design determinants on office building façade for cooling loads and thermal comfort in hot-humid climates, Building and Environment, 226 (2022) 109684. https://doi.org/10.1016/j.buildenv.2022.109684
- Badan Standarisasi Nasional, SNI 03-6389-2000 Konservasi energi selubung bangunan pada bangunan gedung. 1., Badan Standarisasi Nasional: Jakarta, Indonesia, 2020, pp. 1-39.
- US Green Building Council, LEED v4.1 Quality Views, 2020. [Online]. Available: https://www.usgbc.org/credits/core-and-shell/v4-draft/eqc6. [Accessed: 01-Feb-2024].
- GBCI, Perangkat penilaian GREENSHIP (GREENSHIP rating tools), Greensh. New Build. Versi 1.2, Green Building Council Indonesia, Jakarta, Indonesia, 2013, pp. 1-15.
- H. Ohly, M.P. White, B.W. Wheeler, A. Bether, O.C. Ukoumunne, V. Nikolaou, R. Garside, Attention Restoration Theory: A systematic review of the attention restoration potential of exposure to natural environments, Journal of Toxicology and Environmental Health - Part B: Critical Reviews, 19:7 (2016) 305-343. https://doi.org/10.1080/10937404.2016.1196155
- Y. Liu, J. Zhang, C. Liu, Y. Yang, A Review of Attention Restoration Theory: Implications for Designing Restorative Environments, Sustainability, 16:9 (2024) 1-18. https://doi.org/10.3390/su16093639
- M. Keener, The Effects of Nature Exposure on Attention, Department of Psychology of Marietta College: Ohio, USA, 2016.
- J. Moore, Mentally restorative areas for students: impacts of nature on psychophysiological state, University of Tennessee: Chattanooga, USA, 2021.
- J. Baur, Campus community gardens and student health: A case study of a campus garden and student well-being, Journal of American College Health, 70:2 (2022) 377-384. https://doi.org/10.1080/07448481.2020.1751174
- A. Angelidis, W. van der Does, L. Schakel, P. Putman, Frontal EEG Theta/beta ratio as an electrophysiological marker for attentional control and its test-retest reliability, Biological Psychology, 121 (2016) 49-52. https://doi.org/10.1016/j.biopsycho.2016.09.008
- H. Wei, L. Chang, Q. Huang, R. Zhou, Relation between spontaneous electroencephalographic Theta/beta power ratio and test anxiety, Neuroscience Letters, 737 (2020) 135323. https://doi.org/10.1016/j.neulet.2020.135323
- M. Waczynska, N. Sokol, J. Martyniuk-Peczek, Computational and experimental evaluation of view out according to European Standard EN17037, Building and Environment, 188 (2021) 107414. https://doi.org/10.1016/j.buildenv.2020.107414
- M. Kuhlenengel, C.E. Waters, I. Konstantzos, Assessing the impact of outside view on learning: A close look to 17037 'view out' practices through the analysis of 220 classrooms, Journal of Physics: Conference Series, 1343:1 (2019). https://doi.org/10.1088/1742-6596/1343/1/012159
- F. Abd-Alhamid, M. Kent, Y. Wu, Assessment of Window Size and Layout Impact on a View Quality Perception in a Virtual Reality Environment, LEUKOS - Journal of Illuminating Engineering Society of North America, 20:3 (2024). https://doi.org/10.1080/15502724.2023.2262148
- M. Newman, B. Gatersleben, K.J. Wyles, E. Ratcliffe, The use of Virtual Reality in environment experiences and the importance of realism, Journal of Environmental Psychology, 79 (2022) 101733. https://doi.org/10.1016/j.jenvp.2021.101733
- A. Felnhofer, O.D. Kothgassner, M. Schmidt, A-K. Heinzle, L. Beutl, H. Hlavacs, I. Kryspin-Exner, Is Virtual Reality emotionally arousing? Investigating five emotion inducing virtual park scenarios, International Journal of Human Computer Studies, 82 (2015) 48-56. https://doi.org/10.1016/j.ijhcs.2015.05.004
- T.D. Parsons, Virtual Reality for enhanced ecological validity and experimental control in the clinical, affective and social neurosciences, Frontiers in Human Neuroscience, 9:December (2015) 1-19. https://doi.org/10.3389/fnhum.2015.00660
- D.J. Harris, J.M. Bird, P.A. Smart, M.R. Wilson, S.J. Vine, A Framework for the Testing and Validation of Simulated Environments in Experimentation and Training, Frontiers in Psychology, 11 (2020) 1-10. https://doi.org/10.3389/fpsyg.2020.00605
- K. Chamilothori, J. Wienold, M. Andersen, Adequacy of Immersive Virtual Reality for the Perception of Daylit Spaces: Comparison of Real and Virtual Environments, LEUKOS - Journal of Illuminating Engineering Society of North America, 15 (2019) 203-226. https://doi.org/10.1080/15502724.2017.1404918
- K. Yin, E-L. Hsiang, J. Zou, Y. Li, Z. Yang, Q. Yang, P-C Lai, C-L Lin, S-T Wu, Advanced liquid crystal devices for Augmented Reality and Virtual Reality displays: principles and applications, Light: Science and Applications, 11:1 (2022). https://doi.org/10.1038/s41377-022-00851-3
- K.W. May, J. Walsh, R.T. Smith, N. Gu, B.H. Thomas, VRGlare: A Virtual Reality Lighting Performance Simulator for real-time Three-Dimensional Glare Simulation and Analysis, Proc. 37th Int. Symp. Autom. Robot. Constr. ISARC 2020 From Demonstr. to Pract. Use - To New Stage Constr. Robot, no. Isarc, Kitakyushu, Japan, 27-28 October 2020 pp. 32-39. https://doi.org/10.22260/ISARC2020/0006
- J. de Chiara, J. Callender, Time Saver: Standards for Building Types, 4th ed. McGraw-Hill Inc.: New York, USA, 2014.
- D.A. Chi, D. Moreno, J. Navarro, Design optimisation of perforated solar façades in order to balance daylighting with thermal performance, Building and Environment, 125 (2017) 383-400. https://doi.org/10.1016/j.buildenv.2017.09.007
- N. Queiroz, F.S. Westphal, F.O. Ruttkay Pereira, A performance-based design validation study on EnergyPlus for daylighting analysis, Building and Environment,183 (2020). https://doi.org/10.1016/j.buildenv.2020.107088
- R.A. Mangkuto, M. Rohmah, A.D. Asri, Design optimisation for window size, orientation, and wall reflectance with regard to various daylight metrics and lighting energy demand: A case study of buildings in the tropics, Applied Energy, 164 (2016) 211-219. https://doi.org/10.1016/j.apenergy.2015.11.046
- S. Vaisi, F. Kharvari, Evaluation of Daylight regulations in buildings using daylight factor analysis method by radiance, Energy for Sustainable Development, 49 (2019) 100-108. https://doi.org/10.1016/j.esd.2019.02.002
- R.T. Arinta, P. Satwiko, R.R. Widjaja, S. Kusrohmaniah, Brain spatial reconciliation through multisensory integration in the impact of pandemic fatigue on workplace, Frontiers in Human Neuroscience, 18 (2024) 1-11. https://doi.org/10.3389/fnhum.2024.1419889
- A.T. Gifford, K. Dwivedi, G. Roig, R.M. Cichy, A large and rich EEG dataset for modeling human visual object recognition, Neuroimage, 264 (2022) 119754. https://doi.org/10.1016/j.neuroimage.2022.119754
- H.N.A. Logemann, M.M. Lansbergen, T.W.D.P. Van Os, K.B.E. Bocker, J.L. Kenemans, The effectiveness of EEG-feedback on attention, impulsivity and EEG: A sham feedback controlled study, Neuroscience Letters, 479:1 (2010) 49-53. https://doi.org/10.1016/j.neulet.2010.05.026
- I. Sturm, S. Dähne, B. Blankertz, G. Curio, Multi-variate EEG analysis as a novel tool to examine brain responses to naturalistic music stimuli, PLoS One, 10:10 (2015) 1-20. https://doi.org/10.1371/journal.pone.0141281
- J.C. Han, C. Zhang, Y-D. Cai, Y-T. Li, Y-X. Shang, Z-H. Chen, G. Yang, J-J. Song, D. Su, K. Bai, J-T. Sun, Y. Liu, N. Liu, Y. Duan, W. Wang, Neuroimaging features for cognitive fatigue and its recovery with VR intervention: An EEG microstates analysis, Brain Research Bulletin, 221 (2025) 111223. https://doi.org/10.1016/j.brainresbull.2025.111223
- D.H. Lee, K.M. Jang, H.K. Lim, Electroencephalography Changes During Cybersickness: Focusing on Delta and Alpha Waves, Brain Topography, 38:3 (2025). https://doi.org/10.1007/s10548-025-01109-y
- T. Vu, A. van der Meulen, D. Heslenfeld, K. Woodcock, S. Han, L. Krabbendam, Neural responses to affective stimuli across culturally similar and dissimilar situations, Culture and Brain, 8:1 (2020) 1-26. https://doi.org/10.1007/s40167-019-00082-1
- A. Murata, J.S. Moser, S. Kitayama, Culture shapes electrocortical responses during emotion suppression, Social Cognitive and Affective Neuroscience, 8:5 (2013) 595-601. https://doi.org/10.1093/scan/nss036
- B. Kraus, K. Liew, S. Kitayama, Y. Uchida, The impact of culture on emotion suppression: Insights from an electrophysiological study of emotion regulation in Japan, Biological Psychology, 187: November (2024) 108767. https://doi.org/10.1016/j.biopsycho.2024.108767
- Z.H. Pugh, S. Choo, J.C. Leshin, K.A. Lindquist, C.S. Nam, Emotion depends on context, culture and their interaction: Evidence from effective connectivity, Social Cognitive and Affective Neuroscience, 17:2 (2022), 206-217. https://doi.org/10.1093/scan/nsab092
- V. Kurbalija, M. Ivanović, M. Radovanović, Z. Geler, W. Dai, W. Zhao, Emotion perception and recognition: An exploration of cultural differences and similarities, Cognitive Systems Research, 52 (2018), 103-116. https://doi.org/10.1016/j.cogsys.2018.06.009
- E. Valentini, S. Halder, D. McInnerney, J. Cooke, I.L. Gyimes, V. Romei, Assessing the specificity of the relationship between brain alpha oscillations and tonic pain, Neuroimage, 255:March (2022), 119143. https://doi.org/10.1016/j.neuroimage.2022.119143
- S. Huang, J. Qi, W. Li, J. Dong, C.K. van den Bosch, The contribution to stress recovery and attention restoration potential of exposure to urban green spaces in low-density residential areas, International Journal of Environmental Research and Public Health, 18:16 (2021). https://doi.org/10.3390/ijerph18168713
- A. Olszewska-Guizzo, A. Sia, A. Fogel, R. Ho, Features of urban green spaces associated with positive emotions, mindfulness and relaxation, Scientific Reports, 12:1 (2022), 1-13. https://doi.org/10.1038/s41598-022-24637-0
- R. Berto, The role of nature in coping with psycho-physiological stress: A literature review on restorativeness, Behavioral Sciences, 4:4 (2014), 394-409. https://doi.org/10.3390/bs4040394
- E.E. Neri, Challenging Assumptions: Exploring the Extent of Biophilic Design in Interior Environments on Attention Restoration," Florida State University: Florida, USA, 2023.
- D. Dillon, J. Cai, Virtual Reality Greenspaces: Does Level of Immersion Affect Directed Attention Restoration in VR Environments?, J: Multidisciplinary Scientific Journal, 5:3 (2022) 334-357. https://doi.org/10.3390/j5030023
2383-8701/© 2025 The Author(s). Published by solarlits.com. This is an open access article distributed under the terms and conditions of the Creative Commons Attribution 4.0 License.

3029
Total views
Citations
SHARE ON



